Envoltura membranosa de los virus
La cubierta exterior de un virus puede ser una cápside proteica o una envoltura membranosa:
Inicialmente se pensó que la cubierta externa de los virus estaba formada
por un sólo tipo de molécula proteica. Se creía que las infecciones víricas
empezaban por la separación del cromosoma vírico (su ácido nucleico) de la
cubierta proteica, y que a continuación se producía la replicación del
cromosoma dentro de la célula huésped, generándose muchas copias idénticas.
Después de la síntesis de nuevas copias de la envoltura proteica específica del
virus sobre moléculas de RNA mensajero codificadas por el virus, podía
producirse la formación de partículas víricas hijas mediante el ensamblaje
espontáneo de esta cubierta proteica rodeando los cromosomas víricos hijos
(Figura 6-71).
|
Ahora se sabe que estas ideas sobresimplifican extraordinariamente la diversidad
de ciclos vitales víricos que existen. Por ejemplo, la proteína de cubierta
que rodea el ácido nucleico de la mayoría de los virus (la cápside) contiene
más de un tipo de cadena polipeptídica, a menudo dispuesta en varias capas.
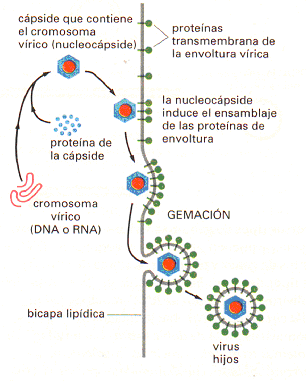
En muchos virus, además, la cápside proteica está recubierta a su vez por una
membrana formada por una bicapa lipídica que contiene proteínas. Muchos de
estos virus recubiertos adquieren su envoltura durante el proceso de brote de
la membrana plasmática (Figura 01). Este proceso de gemación permite a las
partículas víricas abandonar la célula sin romper la membrana plasmática y,
por lo tanto, sin matar a la célula. Los virus
utilizan la maquinaria de tráfico intracelular de sus células huéspedes Todos los virus tienen una cantidad limitada de
ácido nucleico en su genoma, por lo que han de parasitar procesos de la
célula huésped para la mayoría de las etapas de su producción. De hecho,
debido a que habitualmente los productos víricos se sintetizan en grandes
cantidades durante la infección y debido también a que durante su ciclo
vital los virus siguen una rota secuencial a través de los compartimientos de
la célula huésped, las células infectadas por virus se han utilizado como
importantes modelos para trazar las rutas de |
Fig. 01 |
|
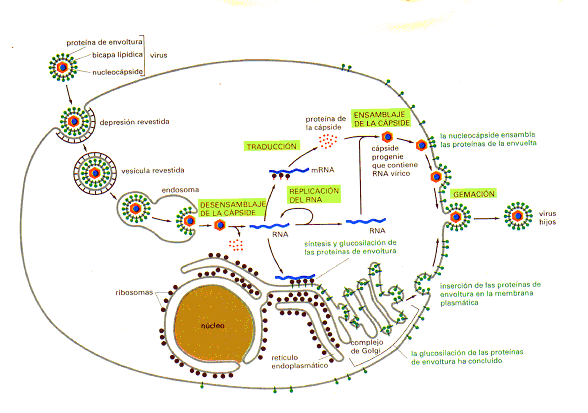
Fig. 02 Ciclo vital
del virus Semliki forest. El virus parasita la célula huésped para la mayor
parte de sus procesos biosintéticos. |
transporte intracelular y para estudiar qué reacciones biosintéticas esenciales
están comparomentalizadas en las células eucariotas. Los virus recubiertos que afectan a las células animales, en los cuales
el genoma está incluido en una membrana de bicapa lipídica, han utilizado la
compartimentalización de la célula hasta un grado especialmente preciso.
Seguir el ciclo vital de un virus recubierto es hacer un "tour" a
través de la célula. Un ejemplo bien estudiado es el virus Semliki forest,
que consiste en un genoma de cadena sencilla de ARN rodeado por una cápside
formado por una envoltura eicosaédrica (20 caras) dispuesta de forma regular
y compuesta por muchas copias de una proteína (denominada proteína C). La
nucleocápside (genoma + cápside) está rodeada por una bicapa lipídica situada
muy cerca, que contiene únicamente tres tipos de cadenas polipeptídicas,
codificadas por el ARN vírico. Estas proteínas de envoltura forman
heterotrímeros situados en la bicapa lipídica y que
interactúan con la proteína C de la nucleocápside, uniendo la membrana con la
nucleocápside. Las zonas glucosiladas de las proteínas de envoltura siempre
están situadas en el exterior de la hicapa lipídica, y cada trímero forma una "espina "
que al |
microscopio electrónico se ve sobresaliendo de la superficie del virus.
La infección se inicia cuando una proteína de envoltura del virus se une a
una proteína de una célula normal que actúa como su receptor en la membrana
plasmática de la célula huésped. Entonces el virus utiliza el proceso
endocítico normal de la célula para entrar en ella mediante endocitosis mediada
por receptor, y es liberado a los endosomas vecinos. Sin embargo, en lugar de
ser transferido desde los endosomas a los lisosomas, el virus se escapa de los
endosomas gracias a las propiedades especiales de una de sus proteínas de
envoltura. Al pH ácido del endosoma esta proteína hace que la envoltura vírica
se fusione con la membrana del endosoma, liberando la nucleocápside desnuda en
el citoplasma. La nucleocápside pierde su cubierta en el citoplasma, liberando
el ARN vírico, que entonces es traducido por los ribosomas de la célula huésped
produciendo una ARN polimerasa codificada por el virus. Esta enzima, a su vez,
produce muchas copias del ARN vírico, algunas de las cuales actúan como
moléculas de ARNm dirigiendo la síntesis de proteínas estructurales del
virus-la proteína C de la cápside y las tres proteínas de la envoltura.
Las proteínas de la cápside y de la envoltura, recién sintetizadas, siguen
vías separadas a través del citoplasma. Las proteínas de la envoltura, como las
proteínas de la membrana plasmática de la célula huésped, son sintetizadas en
los ribosomas que se hallan unidos al ER rugoso; por el contrario, la proteína
de la cápside, como las proteínas citosólicas de la célula, se sintetiza por ribosomas
que no se hallan urudos a membrana. Las proteínas recién sintetizadas de la
cápside se unen al RNA vírico recientemente replicado, formando nuevas
nucleocápsides. Las proteínas de la envoltura, por el contrario, son insertadas
en la membrana del ER, donde son glucosiladas, transportadas hasta el complejo
de Golgi y liberadas a la membrana plasmática (Figura 02).
Finalmente las nucleocápsides víricas y las proteínas de la envoltura se encuentran
en la membrana plasmática. Como resultado de una interacción específica con un
grupo de proteínas de envoltura, la nucleocápside forma una gema cuya envoltura
contiene proteínas de envoltura embebidas en lípidos de la célula huésped. Por
último, la gema se libera, de forma que aparece un nuevo virus independiente de
la célula. E1 agrupamiento de las proteínas de la envoltura mientras se
ensamblan alrededor de la nucleocápside durante la gemación vírica hace que
las proteínas plasmáticas de la célula huésped queden excluidas de la partícula
vírica final.